
合成生物学经典通路系列 · 第四篇
第三篇我们走完了细胞的"基础设施"——糖类、脂质、氨基酸、核苷酸、辅因子。这一篇,我们登上第二层楼,看看微生物如何用初级代谢的"半成品",组装出结构最复杂、价值最高的天然产物。
? 合成生物学经典通路系列
微生物细胞里的"十号线":一张图看懂合成生物学的代谢全景 走进"中央厨房":EMP、TCA、PPP三条核心代谢途径详解 细胞的"基础设施":五大初级代谢产物合成通路全解析 微生物的"奢侈品生产线":五大次级代谢产物合成通路全解析(本文) 配套系统:NADPH、ATP与氧还平衡的设计哲学(待更新)
一、写在前面
第三篇"初级代谢"中,我们看到葡萄糖如何经过中心碳代谢和初级合成通路,变成维持生命的基本物质。这一篇进入次级代谢产物——它们不是生长所必需的,但往往是自然界中结构最复杂、生物活性最强、市场价值最高的化合物。
青蒿素拯救了数百万疟疾患者,紫杉醇是最重要的抗癌药物之一,小檗碱是中药黄连的核心成分,红景天苷正是灵知生物的主打产品。这些产物有一个共同特征:植物中含量极低,化学全合成成本极高,微生物异源合成是突破供给瓶颈的最佳路径。
次级代谢五大类产物各有明确的"构建块"来源——这正是第一篇中建立的分类框架。乙酰CoA是萜类和聚酮的构建块,氨基酸是生物碱和非核糖体肽的构建块,莽草酸途径产生的芳香族氨基酸是酚类/苯丙烷类的构建块。本篇将沿着这三大构建块路线逐一展开。
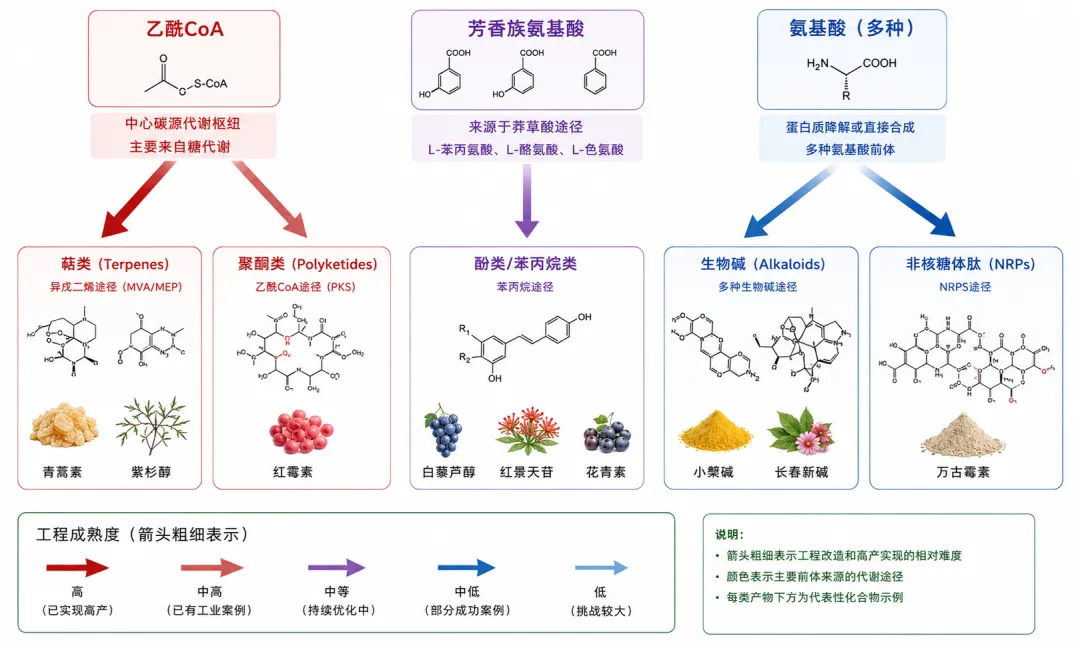
二、萜类/类异戊二烯:自然界最大的分子家族
2.1 五碳积木的拼接逻辑
萜类(terpenoids,也叫类异戊二烯 isoprenoids)是自然界中结构最多样的化合物家族,已知超过80,000种。但不管结构多么复杂,它们都遵循一个简单的规则:全部以五碳的异戊烯基焦磷酸(IPP)及其异构体二甲基烯丙基焦磷酸(DMAPP)为基本构建单元,像搭积木一样拼接而成。
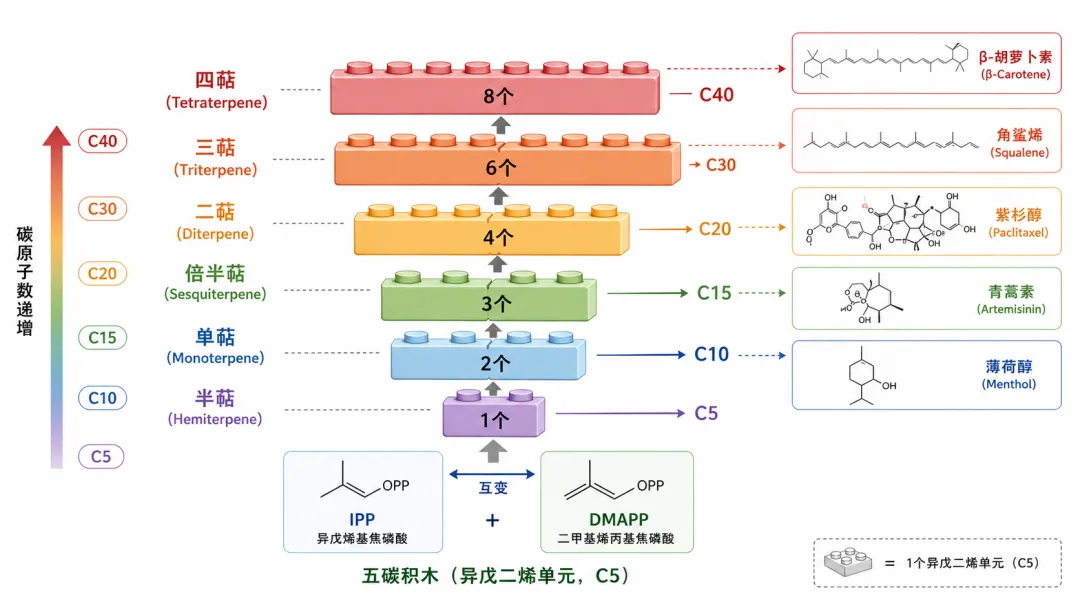
拼接规则:DMAPP + IPP → GPP(C₁₀,单萜前体)→ FPP(C₁₅,倍半萜前体)→ GGPP(C₂₀,二萜前体)。两个FPP头对头缩合生成角鲨烯(C₃₀,三萜前体),两个GGPP缩合生成八氢番茄红素(C₄₀,四萜前体)。
工程成熟度:★★★★☆(四星) — 青蒿酸25 g/L、β-胡萝卜素数g/L,是次级代谢中最成熟的类别
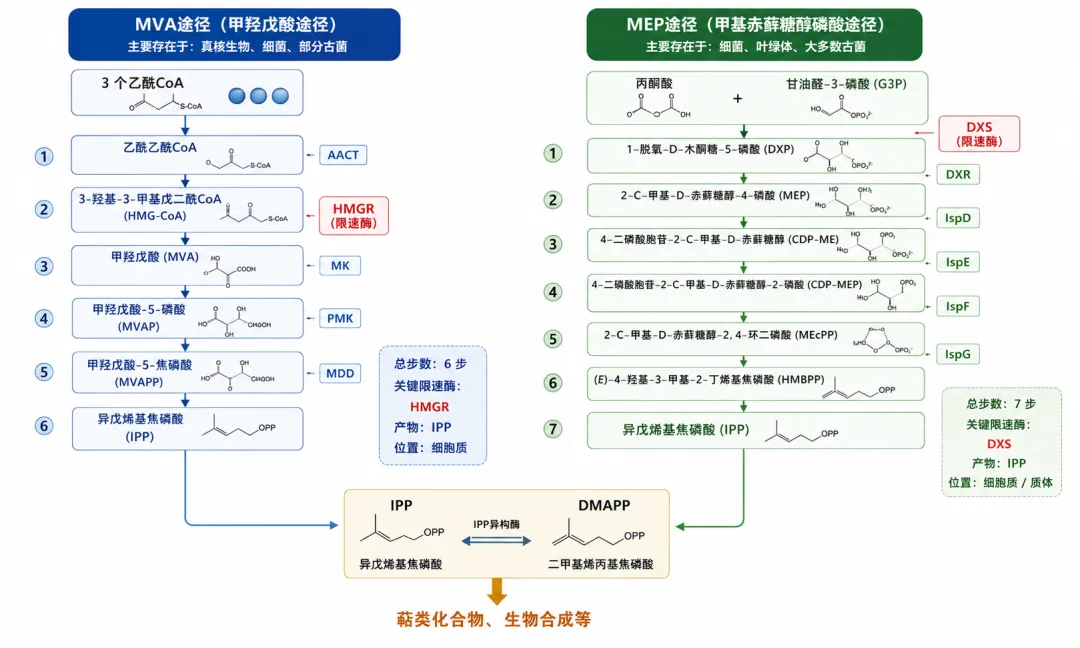
2.2 MVA途径——酵母和动物的"经典路线"
MVA途径(甲羟戊酸途径,Mevalonate pathway)共6步反应,以3分子乙酰CoA为起点:
- 步骤①
乙酰CoA + 乙酰乙酰CoA → 硫解酶(Erg10) → HMG-CoA前体 - 步骤②
HMG-CoA前体 + 第3个乙酰CoA → HMG-CoA合酶(Erg13) → HMG-CoA - 步骤③
HMG-CoA → HMG-CoA还原酶(HMGR) → 甲羟戊酸(MVA)★★★核心限速酶 — 消耗2个NADPH,是整个途径的瓶颈 - 步骤④
MVA → 甲羟戊酸激酶(MK) → 甲羟戊酸-5-磷酸 - 步骤⑤
甲羟戊酸-5-磷酸 → 磷酸甲羟戊酸激酶(PMK) → 甲羟戊酸-5-焦磷酸 - 步骤⑥
甲羟戊酸-5-焦磷酸 → 甲羟戊酸焦磷酸脱羧酶(MVD) → IPP
化学计量:3乙酰CoA + 3 ATP(三磷酸腺苷)+ 2 NADPH(还原型烟酰胺腺嘌呤二核苷酸磷酸酯)→ 1 IPP + 1 CO₂
最经典的工程策略:截短HMGR(tHMGR)。 HMGR的N端有一个约500个氨基酸的跨膜结构域,使酶锚定在内质网膜上,并容易被泛素化降解。截去N端只保留C端约400个氨基酸的催化结构域后,HMGR变成可溶性蛋白,表达量和活性大幅提升。这一改造源自酵母甾醇调控的基础研究,是萜类合成工程中最经典、最广泛使用的策略之一。
2.3 MEP途径——细菌的"替代路线"
MEP途径(甲基赤藓糖醇磷酸途径,Methylerythritol phosphate pathway)共7步反应,以丙酮酸和3-磷酸甘油酸(G3P)为起点:
- 步骤①
丙酮酸 + G3P → DXS(1-脱氧-D-木酮糖-5-磷酸合酶) → DXP(C₅)★★最大瓶颈 — 需要TPP辅酶,释放CO₂ - 步骤②
DXP → DXR(1-脱氧-D-木酮糖-5-磷酸还原异构酶,也叫IspC) → MEP★限速酶 — 消耗NADPH,是抗疟药磷胺霉素的靶点 - 步骤③
MEP → IspD → CDP-ME - 步骤④
CDP-ME → IspE → CDP-ME2P - 步骤⑤
CDP-ME2P → IspF → MEcPP - 步骤⑥
MEcPP → IspG → HMBPP(需要[4Fe-4S]簇) - 步骤⑦
HMBPP → IspH → IPP/DMAPP(比例约5:1)
化学计量:丙酮酸 + G3P + 1 CTP + 1 ATP(三磷酸腺苷)+ 3 NADPH(还原型烟酰胺腺嘌呤二核苷酸磷酸酯)→ IPP/DMAPP
MEP途径存在于大多数细菌和植物叶绿体中。大肠杆菌(E. coli)天然拥有完整的MEP途径,这是E. coli在萜类生产上的天然优势。
2.4 两条途径的对比与组合策略
一个重要策略是在E. coli中异源引入完整的MVA途径。 Jay Keasling团队率先实现了这一点——使E. coli同时拥有MEP和MVA两条IPP合成通路。由于两条途径使用不同的前体(MVA用乙酰CoA,MEP用丙酮酸+G3P),它们可以互补而不互相干扰,萜类产量实现了数量级提升。
2.5 经典案例:青蒿素——从植物到酵母的二十年长征
青蒿素是中国科学家屠呦呦从黄花蒿中发现的抗疟疾药物,她因此获得2015年诺贝尔生理学或医学奖。青蒿素在植物中的含量极低(干重<1%),且化学全合成步骤多、收率低,供给长期不足。
2006年,Ro等在Nature上发表了里程碑式的论文——在酵母中实现了青蒿酸(青蒿素的前体)的微生物合成。核心改造策略是萜类工程的标准范式:
- 过表达tHMGR
→ MVA通量大幅提升 - 上调ERG20
→ FPP合酶过表达,增加FPP供给 - 敲弱ERG9
→ 角鲨烯合酶弱化,阻止FPP被分流到甾醇合成 - 引入三个外源基因
→ 青蒿的ADS(紫穗槐二烯合酶)+ CYP71AV1(细胞色素P450)+ CPR(P450还原酶) - 两阶段发酵
→ 好氧生长期积累生物量 + 限制氧产素期积累产物
酵母内源MVA途径 → FPP → ADS → 紫穗槐-4,11-二烯 → CYP71AV1/CPR → 青蒿醇 → 青蒿醛 → 青蒿酸
青蒿酸经化学半合成 → 双氢青蒿酸 → 青蒿素
这套策略后来被Sanofi放大到工业规模,青蒿酸产量达25 g/L,彻底改变了全球青蒿素的供给格局。
参考文献:Ro DK, et al. (2006) Nature 440:940. PMID: 16612385 | Liao P, et al. (2016) Biotechnol Adv 34:697. PMID: 26995109 | Navale GR, et al. (2021) Appl Microbiol Biotechnol 105:435. PMID: 33394155
三、酚类与苯丙烷类:从氨基酸到植物化学武器
3.1 莽草酸途径:芳香族化合物的唯一入口
如果说萜类的构建块是乙酰CoA,那酚类和苯丙烷类的构建块则完全不同——它们来自莽草酸途径产生的芳香族氨基酸(苯丙氨酸、酪氨酸、色氨酸)。
莽草酸途径(Shikimate pathway)是从磷酸烯醇式丙酮酸(PEP)和赤藓糖-4-磷酸(E4P)到芳香族氨基酸的唯一通路,在所有已知生物中没有替代方案。这意味着,所有芳香族化合物——从苯丙氨酸到黄酮、从色氨酸到吲哚生物碱——都必须经过这条途径:
- 步骤①
PEP + E4P → DAHP合酶(AroG/AroF/AroH) → DAHP★★★绝对限速酶 — 三个同工酶分别受苯丙氨酸/酪氨酸/色氨酸反馈抑制 - 步骤②-③
DAHP → DHQ → DHS - 步骤④
DHS → 莽草酸脱氢酶(AroE) → 莽草酸(Shikimate)消耗NADPH。莽草酸是抗流感药达菲(Tamiflu,奥司他韦)的合成起始原料 - 步骤⑤
莽草酸 → 莽草酸激酶(AroL/AroK) → 莽草酸-3-磷酸 - 步骤⑥
莽草酸-3-磷酸 + PEP → EPSP合酶(AroA) → EPSP草甘膦(Glyphosate,即农达Roundup)的靶点! - 步骤⑦
EPSP → 分支酸合酶(AroC) → 分支酸(Chorismate)三种芳香族氨基酸的分支点
化学计量:2 PEP + 1 E4P + 1 ATP(三磷酸腺苷)+ 1 NADPH(还原型烟酰胺腺嘌呤二核苷酸磷酸酯)→ 1 分支酸
工程改造的核心:使用反馈抗性DAHP合酶突变体(aroGfbr 或 aroFfbr)解除终产物对限速酶的抑制;通过敲除PTS系统和过表达tktA来增加PEP和E4P的供给。
3.2 从分支酸到苯丙烷下游:红景天苷的完整路线
分支酸是三条路线的起点——苯丙氨酸、酪氨酸、色氨酸各走一条分支。对灵知生物而言,酪氨酸分支最重要,因为它是红景天苷和酚类产物的直接前体:
分支酸 → 预苯酸 → 酪氨酸(Tyr)
酪氨酸 → TAL(酪氨酸解氨酶) → 对香豆酸(p-Coumaric acid)
对香豆酸 → 4CL(+CoA(辅酶A), ATP(三磷酸腺苷)) → 对香豆酰CoA
对香豆酰CoA面临三条分支:
分支A(黄酮路线): 对香豆酰CoA + 3×丙二酰CoA → CHS(查尔酮合酶) → 柚皮素查尔酮 → CHI → 柚皮素 → [F3H → F3'H → DFR → ANS → 3GT] → 花青素
分支B(芪类路线): 对香豆酰CoA + 3×丙二酰CoA → STS(芪合酶) → 白藜芦醇(Resveratrol)
分支C(红景天苷路线): 对香豆酸 → … → 酪醇 → UGT(糖基转移酶) + UDP-葡萄糖 → 红景天苷(Salidroside)
白藜芦醇工程是苯丙烷类另一个成功案例。Yuan等(2020)使用E. coli-S. cerevisiae共培养体系——E. coli负责上游(TAL + 4CL),酵母负责下游(STS),利用两个物种各自的代谢优势分担通路负荷。白藜芦醇在E. coli中产量已超过2 g/L。
参考文献:Rodriguez A, et al. (2014) Microb Cell Fact 13:126. PMID: 25200799 | Nabavi SM, et al. (2020) Biotechnol Adv 38:107132. PMID: 30458225
四、生物碱:含氮的"药库"——也是工程难度最大的家族
4.1 为什么生物碱如此难做?
在十类产物中,生物碱的工程成熟度最低(二星),但市场价值极高——长春新碱(抗癌)、吗啡(镇痛)、小檗碱(抗菌/降血糖)、奎宁(抗疟)都是生物碱。生物碱的工程化面临三个独特挑战:
- 通路极长。
从氨基酸到终产物通常需要15-30步酶促反应,远超萜类(<10步)和酚类(<10步)。 - P450氧化酶密集。
几乎每个生物碱通路都包含多个植物来源的细胞色素P450(CYP),而P450在大肠杆菌中功能性表达一直是瓶颈。 - 中间体有毒性。
多巴胺、血清素等中间体在高浓度下抑制微生物生长。
4.2 生物碱按氨基酸前体分类
色氨酸 → 吲哚类生物碱
单萜吲哚类(MIA):长春新碱(抗癌)、蛇根碱(降压) 简单吲哚类:褪黑素、血清素 喹啉类:奎宁(抗疟)、喜树碱(抗癌)
酪氨酸/多巴 → 异喹啉类生物碱(BIA)
苄基异喹啉类:(S)-reticuline是中央分支点 原小檗碱类:小檗碱/黄连素 吗啡类:吗啡、可待因 阿朴啡类:木兰碱
鸟氨酸/赖氨酸 → 莨菪烷类
阿托品(散瞳)、东莨菪碱(抗晕)
4.3 BIA通路详解:小檗碱的微生物合成
小檗碱(berberine,黄连素)是中药黄连的主要活性成分,具有抗菌、降血糖和抗炎作用。它的合成路线是BIA(苄基异喹啉生物碱)通路中研究最透彻的:
L-酪氨酸 → TyrDC(酪氨酸脱羧酶) → 多巴胺(Dopamine) 多巴胺 + 4-HPAA(4-羟基苯乙醛)→ NCS(去甲乌药碱合酶) → 去甲乌药碱(Norcoclaurine)★关键步骤 — Pictet-Spengler反应,形成BIA通路的第一个异喹啉骨架 去甲乌药碱 → 6OMT + CNMT + 4'OMT(三个甲基转移酶) → (S)-reticuline★★★BIA通路的中央分支点
(S)-reticuline面临多条下游分支:
分支A(小檗碱): (S)-reticuline → BBE(小檗碱桥酶) → 黄皮碱(Scoulerine)→ [多步氧化/甲基化] → 小檗碱
分支B(吗啡): (S)-reticuline → SalR/salutaridine合酶 → 吗啡通路
分支C(木兰碱): (S)-reticuline → CYP80G2等 → 木兰碱
分支D(血根碱): (S)-reticuline → 其他P450 → 血根碱(Sanguinarine)
(S)-reticuline是BIA通路的核心枢纽——它是多条下游分支的共同前体。在酿酒酵母(S. cerevisiae)中,(S)-reticuline的产量已达4.6 g/L,是一个令人鼓舞的数字。但从(S)-reticuline继续往下走到小檗碱、吗啡、血根碱等终产物,仍多停留在mg/L级别——因为下游步骤密集地使用P450氧化酶,这些植物来源的酶在微生物中的功能表达仍是主要瓶颈。
4.4 MIA通路:长春新碱——更长的路
长春新碱(vincristine)和长春花碱(vinblastine)是从长春花提取的抗癌药物,属于单萜吲哚生物碱(MIA)。MIA通路比BIA更长,且构建块跨越两个来源——色胺来自色氨酸(氨基酸代谢),裂环番木鳖苷(secologanin)来自GPP经secoiridoid通路合成(萜类代谢):
色胺 + 裂环番木鳖苷 → STR(异胡豆苷合酶) → 异胡豆苷(Strictosidine)★关键缩合 异胡豆苷 → SGD(β-葡萄糖苷酶) → …(数十步反应,涉及多个P450和甲基转移酶)… → 文多灵(Vindoline)+ 长春花宁(Catharanthine)→ [两者偶联] → 长春新碱 / 长春花碱
长春新碱的市场价值巨大(全球约2亿美元/年),因为植物提取产量极低(约0.0005%干重)且结构复杂无法化学全合成。但完整的MIA微生物合成目前仍是合成生物学的前沿挑战——通路太长、P450太多。
4.5 生物碱工程的共同策略
面对这些挑战,代谢工程师发展了几种通用策略:
- 使用酵母底盘
——酵母的内质网为P450提供了天然的功能环境(内质网膜上的CPR为P450提供电子)。 - 分步引入
——不追求一次完成整条通路,而是先构建核心片段(如酪氨酸→reticuline),验证可行性后再向下延伸。 - 区室化表达
——将有毒中间体的合成限制在特定细胞器(如过氧化物酶体),减轻对细胞整体的毒害。
参考文献:Cao L, et al. (2024) ACS Synth Biol 13:4017. PMID: 39579377 | Mistry V, et al. (2022) Appl Microbiol Biotechnol 106:2083. PMID: 35333954 | Yao L, et al. (2023) Biotechnol Adv 72:108087. PMID: 37722606
五、聚酮(PKS,聚酮合酶):自然界的分子装配线
5.1 聚酮与脂肪酸的"亲缘关系"
聚酮合酶(PKS)与脂肪酸合酶(FAS)在反应化学上同源——都基于Claisen缩合,都以丙二酰CoA为延伸单元。但PKS比FAS"灵活"得多:FAS每一轮都执行完整的"缩合→还原→脱水→还原"四步反应,产生全饱和的碳链;而PKS可以选择性地跳过某些还原步骤,保留酮基、羟基或双键,从而产生结构多样性远超脂肪酸的产物。
PKS分为三种类型:
- Ⅰ型PKS
:大型多功能蛋白(~300 kDa/模块),装配线式,代表产物:红霉素、雷帕霉素、洛伐他汀 - Ⅱ型PKS
:离散蛋白质亚基,迭代使用同一套酶,代表产物:四环素、阿霉素 - Ⅲ型PKS
:单一蛋白(~42 kDa),直接用CoA底物,代表产物:查尔酮、白藜芦醇、柚皮素
注意Ⅲ型PKS——查尔酮合酶(CHS)和芪合酶(STS)实际上就是Ⅲ型PKS。这意味着酚类/苯丙烷类中的黄酮和白藜芦醇,从化学本质上看也是聚酮。
5.2 Ⅰ型PKS的模块化逻辑——以红霉素PKS为例
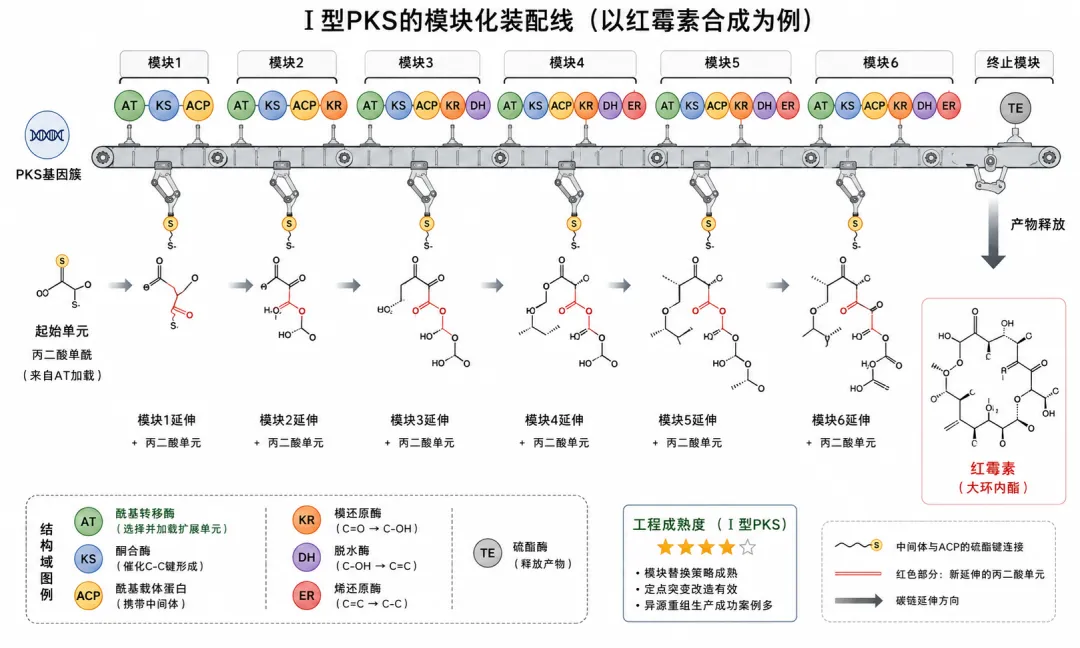
Ⅰ型PKS的工作方式像一条工厂装配线——每个模块负责一轮延伸和可选的修饰:
核心结构域(每个模块必有):
- AT(酰基转移酶)
= "选择"底物 → 丙二酰CoA或甲基丙二酰CoA - KS(酮合酶)
= "延伸"碳链 → Claisen缩合,形成C-C键 - ACP(酰基载体蛋白)
= "运载"中间体 → 磷酸泛酰巯基乙胺"摆臂"共价连接底物
可选修饰结构域:
- KR(酮还原酶)
:-C(=O)- → -CH(OH)- [NADPH] - DH(脱水酶)
:-CH(OH)- → -CH=CH- [-H₂O] - ER(烯还原酶)
:-CH=CH- → -CH₂-CH₂- [NADPH]
终止结构域:
- TE(硫酯酶)
:释放最终产物(常伴随大环内酯化)
红霉素PKS(DEBS)的模块组成:
DEBS1: 模块1(AT→KS→ACP→KR)→ 模块2(AT→KS→ACP→KR→DH→ER)
DEBS2: 模块3(AT→KS→ACP→KR)→ 模块4(AT→KS→ACP→KR→DH→ER)
DEBS3: 模块5(AT→KS→ACP→KR)→ 模块6(AT→KS→ACP→KR→TE)
6个模块 → 14元大环内酯骨架 → 红霉素
每个模块的还原状态决定产物的结构特征:无修饰域则保留β-酮基(-C(=O)-),只有KR则变成β-羟基(-CH(OH)-),KR+DH产生双键(-CH=CH-),KR+DH+ER则完全饱和(-CH₂-CH₂-)。这四种状态就像四进制编码——6个模块的组合决定了最终产物的精确结构。
5.3 PKS的工程策略
模块重组(combinatorial biosynthesis)是PKS工程的核心思路——交换不同PKS的模块来创造自然界不存在的"非天然"聚酮。例如将红霉素PKS的模块3替换为雷帕霉素PKS的模块,就能生成具有新结构的大环内酯。替换AT结构域可以改变底物特异性(甲基丙二酰CoA ↔ 丙二酰CoA),效果是产物多一个或少一个甲基。
但PKS异源表达面临巨大挑战——基因簇通常超过30 kb,在模式菌株中的表达量低,且前体(丙二酰CoA、甲基丙二酰CoA)的供给常常不足。目前,大多数聚酮类抗生素仍由天然产生菌株(Streptomyces等)的工业发酵生产,异源表达主要还处于实验室阶段。
参考文献:Calcott MJ, et al. (2026) Curr Opin Microbiol 80:102547. PMID: 41967291 | Chen B, et al. (2026) Nat Prod Rep 43:475. PMID: 40719200
六、非核糖体肽(NRPS,非核糖体肽合成酶):不用核糖体的肽类工厂
6.1 NRPS与核糖体的根本区别
核糖体按照mRNA的指令装配蛋白质,只能使用20种标准氨基酸,按模板依次连接。NRPS(非核糖体肽合成酶)则完全不同——它不需要mRNA模板,而是按照自身模块的物理结构来"盲装"肽链。每个模块识别、激活并连接一个氨基酸(或非标准氨基酸),还能在装配过程中进行甲基化、氧化、环化等化学修饰。
正因为不受遗传密码约束,NRPS能使用数百种非标准氨基酸作为构件,产生了结构极其多样的产物。
6.2 NRPS的模块化逻辑
核心结构域:
- A(腺苷化域)
= "识别并激活"特定氨基酸 → ATP + AA → AA-AMP → AA-S-PCP - T/PCP(肽酰载体蛋白域)
= "运载"中间体 → "摆臂"在模块间传递正在生长的肽链 - C(缩合域)
= "形成肽键" → 催化上游肽链与下游氨基酸缩合
可选修饰结构域:
- Ep(表异构化域)
:L-氨基酸 → D-氨基酸 - NMT(N-甲基转移酶)
:增加疏水性和代谢稳定性 - Ox(氧化域)
:氧化环化 - R(还原酶域)
:释放时还原酰胺键
终止结构域:
- TE(硫酯酶)
:水解释放 / 环化释放
NRPS遵循"一个模块,一个氨基酸"的基本规则(one module, one amino acid):
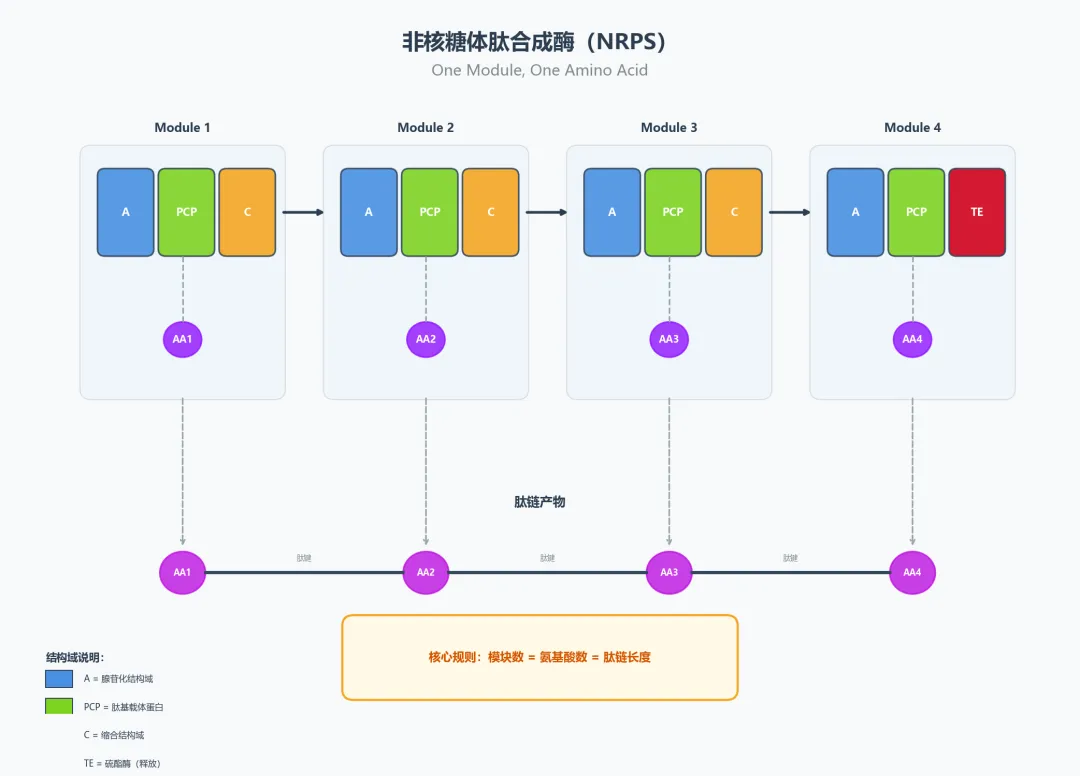
6.3 代表产物
万古霉素(7个氨基酸,Amycolatopsis orientalis)— 革兰阳性菌感染(MRSA最后防线)
环孢菌素A(11个氨基酸,Tolypocladium inflatum)— 免疫抑制剂(器官移植)
短杆菌肽A(15个氨基酸,Bacillus brevis)— 局部抗菌
青霉素/头孢菌素(3个氨基酸,Penicillium / Acremonium)— β-内酰胺抗生素
博来霉素(PKS-NRPS混合,Streptomyces verticillus)— 抗癌
6.4 PKS-NRPS混合系统
许多天然产物由PKS-NRPS杂合装配线合成——同一装配线上既有聚酮延伸模块(使用丙二酰CoA),又有氨基酸添加模块(使用氨基酸)。例如埃博霉素(Epothilone):PKS模块延伸聚酮链 → NRPS模块加入一个氨基酸 → 再由PKS继续延伸 → 最终环化。
这种混合系统产出的化合物结构更加复杂,也提供了更多的工程改造空间——但也意味着更大的异源表达挑战。
2026年,Calcott和Chen等在综述中提出:下一阶段的方向是建立统一的"基因簇—底盘—培养"流水线,用自动化工具加速PKS/NRPS基因簇的设计、底盘适配和发酵优化。这意味着,那些目前仍由天然菌株工业发酵生产的聚酮和非核糖体肽,有望在未来十年内逐步转向工程化底盘的标准化生产。
参考文献:Chen B, et al. (2026) Nat Prod Rep 43:475. PMID: 40719200 | Calcott MJ, et al. (2026) Curr Opin Microbiol 80:102547. PMID: 41967291
写在最后
这篇文章把次级代谢的"五大奢侈品生产线"讲清楚了——萜类用五碳积木拼接出80,000种结构,酚类/苯丙烷类从莽草酸途径走向黄酮和白藜芦醇,生物碱用15-30步反应编织出最复杂的药库,聚酮和自然界的分子装配线以模块化的方式创造无限可能,非核糖体肽则突破了遗传密码的限制。
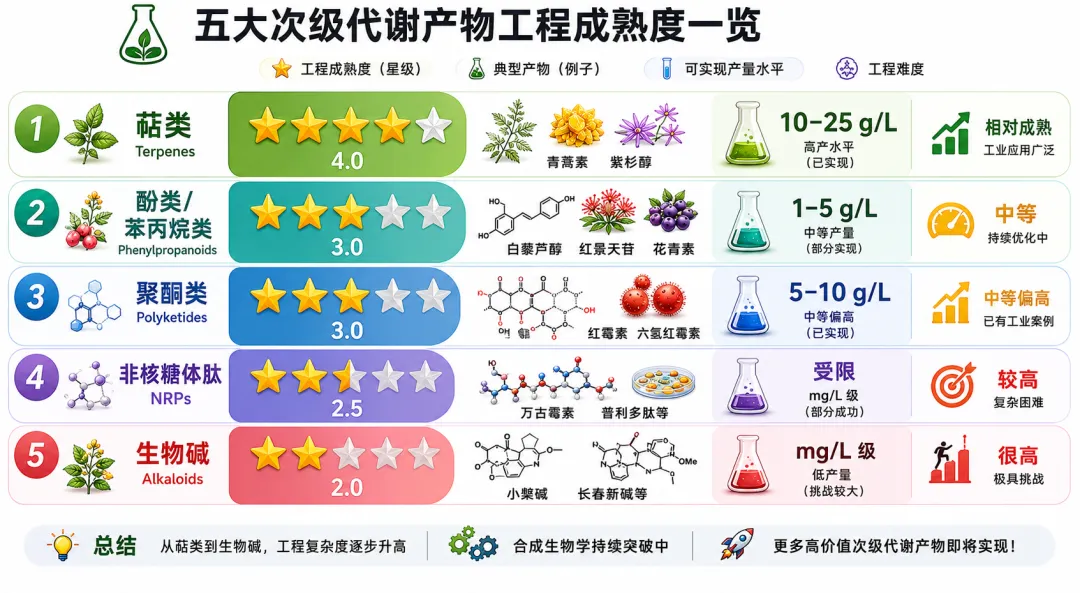
从工程成熟度来看,次级代谢明显落后于初级代谢——萜类(四星)> 酚类/苯丙烷类(三星)> 聚酮(三星)> 非核糖体肽(二星半)> 生物碱(二星)。但这个梯度也揭示了机会:越难做的产物,单位价值越高,技术壁垒越深。
至此,这个系列已经走过了四站:
- 第一篇:
一张全景地图——十类产物、七个枢纽分子、三维平衡框架 - 第二篇:
中心代谢枢纽——EMP、TCA、PPP三条核心途径的详解 - 第三篇:
初级代谢产物——糖类、脂质、氨基酸、核苷酸、辅因子的完整合成通路 - 第四篇:
次级代谢产物——萜类、酚类、生物碱、聚酮、NRPS的工程化策略
第五篇也是最后一篇,我们将回到"基础设施"层面,讨论那些支撑所有产物合成的配套系统——NADPH/ATP的供给与平衡、氧化还原状态的调控、以及如何在代谢网络层面进行系统级优化。这是把"能做"变成"做得起"的关键。
如果你在做任何一个天然产物的生物合成项目,建议收藏这个系列。因为无论你最终的目标产物是什么,碳流都要从同一个起点出发——
葡萄糖。
参考资料
1. Ro DK, et al. (2006) Nature 440:940 — 青蒿素的酵母合成
2. Liao P, et al. (2016) Biotechnol Adv 34:697 — 萜类工程综述
3. Navale GR, et al. (2021) Appl Microbiol Biotechnol 105:435 — 青蒿素发酵优化
4. Rodriguez A, et al. (2014) Microb Cell Fact 13:126 — 苯丙烷类工程
5. Nabavi SM, et al. (2020) Biotechnol Adv 38:107132 — 白藜芦醇工程
6. Cao L, et al. (2024) ACS Synth Biol 13:4017 — 生物碱微生物合成
7. Mistry V, et al. (2022) Appl Microbiol Biotechnol 106:2083 — BIA通路工程
8. Yao L, et al. (2023) Biotechnol Adv 72:108087 — 生物碱合成综述
9. Calcott MJ, et al. (2026) Curr Opin Microbiol 80:102547 — PKS/NRPS工程
10. Chen B, et al. (2026) Nat Prod Rep 43:475 — 聚酮合成酶综述
免责声明
本文仅供学术交流和行业技术科普参考,不构成任何医疗建议或产品功效宣传。文中提及的天然产物生物活性均基于已发表的科研文献,不代表对任何疾病的治疗承诺。具体应用请遵循相关法规和专业指导。
关于本系列
生物研发部知识基座系列,面向合成生物学从业者、原料厂商、生物科研爱好者的深度技术科普。拒绝碎片化,只做系统化。
© 2026 生物研发部 · 合成生物学经典通路系列
